开·云体育app下载安装 痘病毒的形态结构、传播方式及复制周期
痘病毒的形态结构、传播方式及复制周期
在1980年于世界范围内将天花根除之后,人们基本上就不再去开发天花病毒疗法了。2022年世界范围内出现猴痘爆发,这使得天花再度进入公众视野,人们担忧天花病毒或者与它同属的猴痘病毒有可能成为恐怖主义潜在的生物武器。天花病毒也就是Variola virus,猴痘病毒是Monkeypox virus,简称MPXV,它们同属于痘病毒科里的脊索痘病毒亚科中的正痘病毒属。痘病毒是已知的最大的动物病毒,它能够感染大多数的脊椎动物以及无脊椎动物。痘病毒是一种双链 DNA 病毒,它有着约 200 个不同的基因,基因产物的一个子集能够凭借自身独立于宿主细胞核去执行病毒所必需的功能,与此同时其他基因产物能够在广泛的范畴内使用以及调节宿主细胞和免疫系统。
在关乎人类的流行病历史当中,存在着一些相对比较有名的正痘病毒,它们分别有指一种被称作天花病毒的病毒,其标记为Variola virus,还有被叫做痘苗病毒的病毒,它记作Vaccinia virus,另外也叫VACV ,又有所谓名为牛痘病毒的病毒,表示为Cowpox virus,亦称为CPXV ,以及一类被称作猴痘病毒的病毒,记之为Monkeypox virus,亦称MPXV 。将引起人类天花的病原体称为天花(Variola virus),在美国被发现存在两种临床流行病学变种,分别是大天花病毒(Variola major virus)以及小天花病毒(Variola minor virus或alastrim virus),大天花病毒致死率为5 - 40%,小天花病毒致死率为1%,从致死率方面看大天花病毒的毒性更强。牛痘病毒(Cowpox virus,CPXV),因其与奶牛乳头以及挤奶工人手上的脓疱病变存在关联而获得此名称。令人感到有趣的是,在自然界这个范畴当中,牛并非是CPXV的天然宿主,由其引发的牛痘也不是属于那种常见类型的疾病,痘苗病毒也就是(Vaccinia virus,VACV)这样一种在实验室环境里被广泛加以使用的痘病毒模型,它曾经运用在制备天花疫苗这个方面,里面M - 63(苏联)、Lister(英国)、天坛(中国)以及Wyeth、IHD(美国)这样的VACV毒株在使用上是最为广泛的。
痘病毒基因组
痘病毒的基因组kia云手机版登录,是呈线性的双链DNA,其两端有着长度会变化的反向串联重复序列,最终形成发夹结构。就痘苗病毒Copenhagen以及WR病毒株完整的测序信息而言,痘苗病毒基因组全长是191kbp,至于12kbp的反向末端重复序列,也就是ITR,它的末端连接着共价的单链发夹环,该序列富含AT,其中包含短的直接重复序列以及多个ORF。痘病毒能够完全在细胞质里实现复制,对于宿主细胞的DNA以及RNA依赖程度比较低。病毒能够借助产生以及分解大的串联分子,进而合成全新的基因,新基因的包装并不严格,这为基因组的扩增营造了有利的环境条件,在这个过程当中病毒获取到新的功能,最终完成进化。
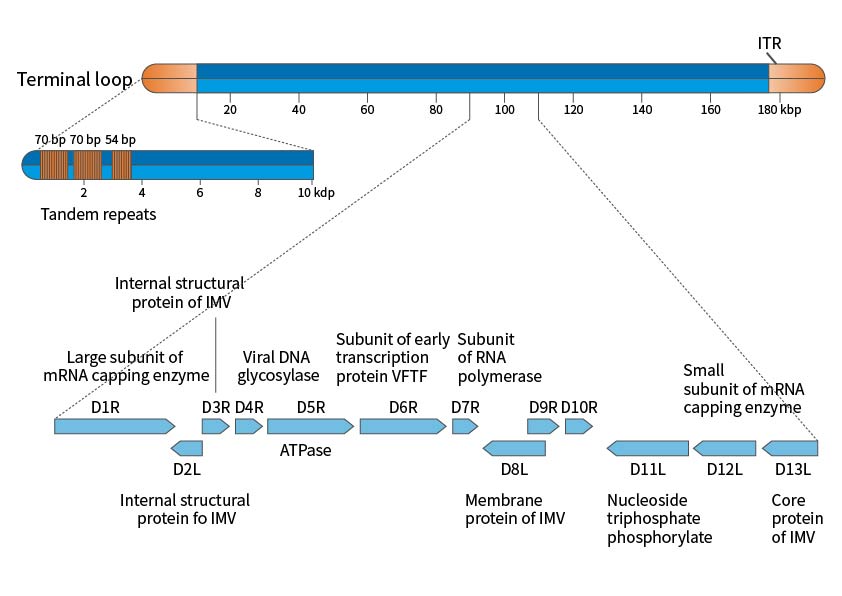
图1. 痘苗病毒基因组示意图
痘病毒形态结构
痘病毒科属于一类病毒,这类病毒是迄今所知结构最为复杂的,痘病毒粒子呈现出椭圆形,或者呈现出砖状的结构,其长度大概是200 - 400nm,轴比为1.2 - 1.7。膜是典型的脂蛋白双分子层,厚度为50 - 55nm,它包裹在核心的周围,外表面布满了随机排列的小管元件,也就是STE,这些小管元件平均宽度是7nm,长度是100nm。由核,和被膜包围着的那些具有足够传染性的相关侧体所组成的病毒粒子,然而,对于某些病毒株以及某些细胞的感染而言,病毒粒子会获取一个内含自身独特化学成分的额外脂质双分子层,此脂质双分子层被称为包膜(Envelop),这种包膜所含的磷脂大概是未包膜病毒粒子所含磷脂的两倍左右,大量研究表明,病毒包膜上的抗原能够诱发免疫反应,从而保护宿主去抵御痘病毒的侵袭,痘病毒包膜之中至少包含着 7 种不一样的糖蛋白,以及一种主要的非税基化酰化多肽。
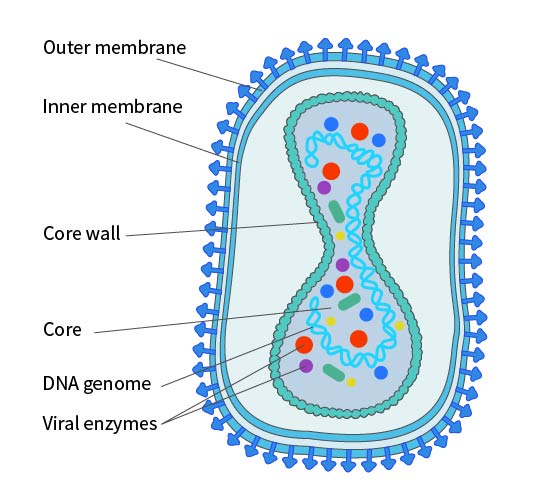
图2. IMV形式的痘病毒结构示意图
注:是否存在内膜尚有争议。
痘病毒传播方式
痘病毒种类不同,宿主不同,病毒的传播过程差别较大,通常有这些方式:在细胞质中保持游离状态,迁移到细胞表面,通过微绒毛排出,被来自高尔基体的两层膜包裹,运输到细胞表面,然后被包膜释放,来自高尔基体的内池膜,通过出芽形成大液泡获得包膜,与无膜的液泡结合,以嗜酸的细胞质结构形式,与一种蛋白质物质相结合,该结构称为a型包涵体(ATI)。
痘苗病毒复制周期
痘病毒的复制周期是依病毒的种类不同而有所差异的,比如说以模型病毒——VACV来讲,复制通常是在感染之后的2到5个小时那里发生。病毒成熟是要历经五个阶段的,分别为未成熟病毒体(IV)、细胞内成熟病毒体(IMV)、细胞内包膜病毒体(IEV)、细胞相关包膜病毒体(CEV)以及细胞外包膜病毒体(EEV)。而IMV和EEV是痘苗病毒的主要传染形式,EEV比起IMAL更具有传染力,它能够通过非pH依赖的方式跟质膜相互融合,进而进到细胞里面。EEV承担着胞间传输的职责,最终呈现为IMV的形态从细胞释放至外部环境里,相较于EEV而言IMV更为稳定,其肩负着宿主间传输的任务。
脱壳
病毒与细胞膜互相融合之际,出现初级脱壳现象kiayun手机版登录入口,病毒的核心被释放至细胞质当中。核心之内不只有病毒基因组,还存在依赖于病毒DNA故而产生的RNA聚合酶,有特异性用以识别病毒早期基因启动子所需要的“起始”蛋白质,更兼有几种能够修改病毒转录本的RNA处理酶。核心被释放以后便合成病毒早期mRNA(见图3,处于步骤2之时),这些mRNA彰显出细胞mRNA的典型特性,在被细胞予以识别之后进行翻译(见图3,处于步骤3之时)。大约有一半的病毒基因在感染的早期阶段得以表达。细胞中分泌出一些早期蛋白,这些早期蛋白与细胞生长因子具备序列相似性(图3,步骤4),它们诱导邻近细胞增殖,或者诱导宿主的抗免疫防御机制。早期蛋白质的合成促使第二次脱壳,核心壁得以打开,含基因组的核蛋白复合体从核心处释放出来(图3,步骤5),病毒早期基因表达就此停止。
复制
病毒DNA基因组复制由早期蛋白催化,此为图3中的步骤6 ,新合成的病毒DNA分子充当了下个复制周期的模板,这是图3中的步骤7 ,它们还是病毒中间期基因的转录模板,即图3 中的步骤8 。中间基因转录激活需特定病毒蛋白(早期基因的产物) 将病毒RNA聚合酶上的中间启动子特异性赋予 ,还有从受感染细胞核重新定位到细胞质的宿主细胞蛋白(Vitf2) 。位于中间的mRNA所编码的蛋白质,此蛋白质在图3的步骤9中有所体现,它包含着晚期基因转录所需要的蛋白,该蛋白在图3的步骤10中呈现。末期基因编码用于构建病毒粒子的蛋白质,还编码病毒粒子的酶以及其他组装过程所需的早期起始蛋白。这些病毒蛋白是借助细胞翻译机制来合成的,此合成过程在图3的步骤11里展示。
组装
无糖基化的乃是病毒膜蛋白,于组装早期细胞膜所起作用存有争议(图3,步骤12),最初进行组装从而形成未成熟病毒粒子(图3,步骤13),此为一种由膜分隔开来的球形粒子,能够从细胞分泌途径的早期获取到,该病毒颗粒成熟之后会形成砖状IMV(图3,步骤14),在细胞裂解之际予以释放(图3,步骤15),另外,颗粒能够从跨高尔基体或者早期内小体室获取第二层双层膜,借此形成胞内包膜病毒粒子(IEV)(图3,步骤16)。
分离
IEV移动至细胞表面的微管那里云手机网页版,跟质膜融合进而形成细胞相关病毒粒子也就是 CEV(呈现于图 3,步骤为 17)。这些 CEV 促使肌动蛋白聚合起来,推动肌动蛋白直接转移至周围细胞(参照图 3,步骤是 步骤为 18);它们还能够从膜上分离出去,从而形成 EEV。
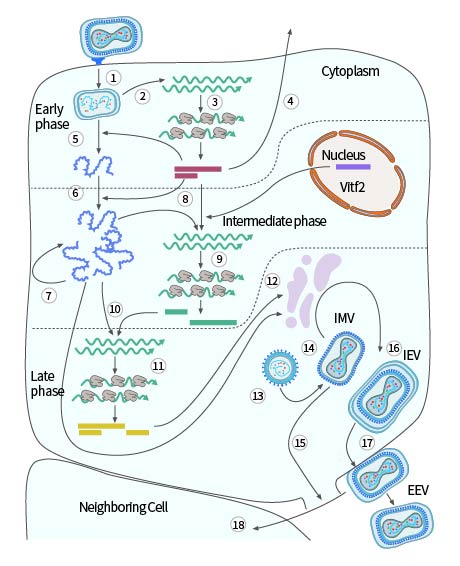
图3. 痘苗病毒的单细胞繁殖周期


